Содержание
Виды коробок передач. Какая лучше
Каждый водитель наверняка помнит слова своего первого инструктора. «Прежде чем повернуть ключ зажигания, убедись, что авто стоит на нейтральной передаче». В данной статье будут рассмотрены виды коробок передач, их отличия друг от друга, преимущества и недостатки, а так же сфера применения.
Усреднено, коленчатый вал двигателей большинства легковых автомобилей имеет рабочие обороты от 800 до 8000 в минуту. При этом, пик мощности приходится на четыре – пять тысяч оборотов. Безусловно, такой диапазон угловых скоростей не отвечает условиям эксплуатации любого колесного транспортного средства в целом и легковых автомобилей в частности.
Основное назначение автомобильной коробки передач – изменение частоты вращения, а так же крутящего момента, передаваемого от коленвала двигателя к ведущим колесам транспортного средства.
Первым узлом такого рода стала механическая коробка передач. Бытует такое мнение, что слово «механическая» попало в название агрегата из за неправильно понятого сокращения, принятого в англоязычной технической литературе. Буквы MT обозначают manual transmission, что означает «ручная, переключаемая рукой передача», а вовсе не mechanical – механическая.
Буквы MT обозначают manual transmission, что означает «ручная, переключаемая рукой передача», а вовсе не mechanical – механическая.
Содержание
- Как это работает
- Четырехступенчатая коробка передач и схема ее работы
- Пятиступенчатая коробка передач
- Несинхронизированые механические коробки передач
- Синхронизированные МКП
- История развития МКП, от Ford T до Bugatti Veyron
- Автоматические и полуавтоматические коробки передач
- Сравнительная характеристика видов коробок передач
Как это работает
Проще всего пояснить принцип действия данного узла на примере работы механической коробки передач. По сути, МКП это многоступенчатый понижающий редуктор, собранный по трехвальной, а реже по двухвальной схеме. Первичный, или ведущий вал посредством сцепления соединен с маховиком ДВС. Вторичный, или ведомый вал имеет жесткое соединение с карданным валом автомобиля. Третий, промежуточный, вал необходим для передачи оборотов от ведущего вала к ведомому. Валы располагаются параллельно друг другу и собраны в едином корпусе.
Валы располагаются параллельно друг другу и собраны в едином корпусе.
На ведущем валу располагается шестерня, передающая движение промежуточному валу. Промежуточный вал оснащен блоком мертво закрепленных шестерен, часто изготовленных как единое целое. Шестерни ведомого вала расположены в щлицах оси или специальных ступицах. Между ними располагаются муфты включения передач, которые вращаются вместе с валом, но способны передвигаться по его продольной оси. Шестерни и муфты ведомого вала могут взаимодействовать друг с другом при помощи зубчатых венцов на своих торцевых поверхностях.
При включении какой либо передачи, кроме заднего хода, муфта, ответственная за ее включение соединяется с соответствующей шестерней и блокирует ее. Двигаясь как единое целое, ведомый вал передает вращение на карданный.
Поступательное движение муфте сообщает водитель транспортного средства, воздействуя на нее при помощи ручки переключения передач, взаимодействующей с вилками и ползунами коробки.
Четырехступенчатая коробка передач и схема ее работы
Цветом выделены:
- Первичный вал – оранжевый
- Вторичный – желтый
- Промежуточный – серый
Цифро-буквенные обозначения указывают на номер передачи и задний ход.
Нейтральное положение и включение первой передачи
Пятиступенчатая коробка передач
Видеоролик, который демонстрирует принцип работы.
Несинхронизированые механические коробки передач
Скорости вращения шестерен ведомого вала значительно отличаются одна от другой. В этом случае, при попытке переключения передачи муфта просто не сможет соединиться с требуемой шестерней, а зубчатый венец будет разрушен. Для приблизительного уравнивания скоростей вращения шестерни и муфты используется прием под названием «двойной выжим». При переключении на более высокую передачу, водитель сначала выжимает сцепление, затем переводит рычаг переключения в нейтральное положение. Промежуточный вал, а следовательно, и ведомый, прекращают вращение. Затем отпускает сцепление, выжимает и включает нужную передачу.
Промежуточный вал, а следовательно, и ведомый, прекращают вращение. Затем отпускает сцепление, выжимает и включает нужную передачу.
При переключении с повышенной передачи на пониженную водителю необходимо провести подобные манипуляции, но в момент, когда коробка выключена, следует нажать на педаль акселератора. Этот прием называется «двойной выжим с перегазовкой».
Несинхронизированные МКП использовались в легковых автомобилях вплоть до 40-х годов двадцатого столетия. Сегодня они используются исключительно в спортивных машинах, и вот по каким причинам:
- Скорость переключения выше, чем у синхронизированных аналогов
- Лучше переносят ударные высокие нагрузки, возникающие в процессе эксплуатации
Синхронизированные МКП
Данные типы коробок передач оснащены дополнительными элементами – синхронизаторами. Шестерни, располагающиеся на ведомом валу, имеют конусную торцевую поверхность. Между каждой шестерней и муфтой переключения находится бронзовое кольцо – синхронизатор. Начиная движение, муфта подхватывает кольцо и прижимает его к торцевой поверхности шестерни. За счет трения, скорости вращения шестерни и муфты уравниваются, после чего происходит их окончательное соединение с помощью зубчатого венца. На сегодняшний день все современные автомобили, на которых стоит 4-х, 5-ти или 6-ти ступенчатая МКП, оснащены синхронизаторами.
Начиная движение, муфта подхватывает кольцо и прижимает его к торцевой поверхности шестерни. За счет трения, скорости вращения шестерни и муфты уравниваются, после чего происходит их окончательное соединение с помощью зубчатого венца. На сегодняшний день все современные автомобили, на которых стоит 4-х, 5-ти или 6-ти ступенчатая МКП, оснащены синхронизаторами.
История развития МКП, от Ford T до Bugatti Veyron
Со времен появления первого автомобиля и по наши дни конструкторы используют следующие типы МКП:
- 2-х ступенчатая планетарная коробка стояла на знаменитом Ford T, чей выпуск начался в 1908 году.
- 3-х ступенчатая МКП появилась в 1920 году. Просуществовала до конца 60-х годов двадцатого века. Встречается на раритетных автомобилях, находящихся в частных коллекциях.
- Четырехступенчатая МКП появилась на свет всего на три года позже трехступенчатой. Но из за отсутствия синхронизаторов плохо выдерживала ударные нагрузки при использовании неопытным водителем.
 Большинство производителей устанавливали на свои автомобили трехступенчатую МКП, четырехступенчатая считалась спортивной опцией.
Большинство производителей устанавливали на свои автомобили трехступенчатую МКП, четырехступенчатая считалась спортивной опцией. - Пятиступенчатая коробка передач появилась в 70-х годах прошлого столетия, а уже через десять лет ей оснащалось подавляющее большинство легковых автомобилей.
- 6-ти, 7-ми, 8-ми ступенчатые коробки разрабатывались с 2000 года, и на данный момент устанавливаются на автомобили Элит класса. К примеру, 5-ти ступенчатая коробка, оснащенная двумя дополнительными передачами Overdrive, стоит на BMW M5.
 Большинство производителей устанавливали на свои автомобили трехступенчатую МКП, четырехступенчатая считалась спортивной опцией.
Большинство производителей устанавливали на свои автомобили трехступенчатую МКП, четырехступенчатая считалась спортивной опцией.Автоматические и полуавтоматические коробки передач
Несмотря на потрясающие возможности современных коробок передач, их конструкция основана на той же, проверенной временем МКП. Изменения затронули привод переключающих муфт и способ передачи крутящего момента от коленвала двигателя на ведущий вал коробки, в остальном же схема осталась неизменной.
Отдельным рядом стоят устройства под названием вариаторы, их принцип действия будет рассмотрен отдельно.
Автоматическая коробка передач или АКПП состоит из гидротрансформатора и обычной 5-ти или 6-ти ступенчатой МКП. Роль гидротрансформатора заключается в плавном выравнивании скоростей вращения ведущего и коленчатого валов. По достижении нужной скорости блокировочная муфта переводит гидротрансформатор в режим гидромуфты. За работу АКПП отвечает электронный блок управления.
Роботизорованная коробка передач представляет собой МКП, в которой функции переключения передач и выключения сцепления полностью автоматизированы. Электронный блок управления и электромеханические сервоприводы справляются с поставленной задачей не хуже профессионального гонщика.
Сравнительная характеристика видов коробок передач
Большинство неофитов, выбирающих свой первый автомобиль, задаются вопросом – какую коробку выбрать? Механику или автомат. А, может, робот? Четырехступенчатая механика не котируется, нужна как минимум, пятиступенчатая коробка передач. Или 7G-Nronic.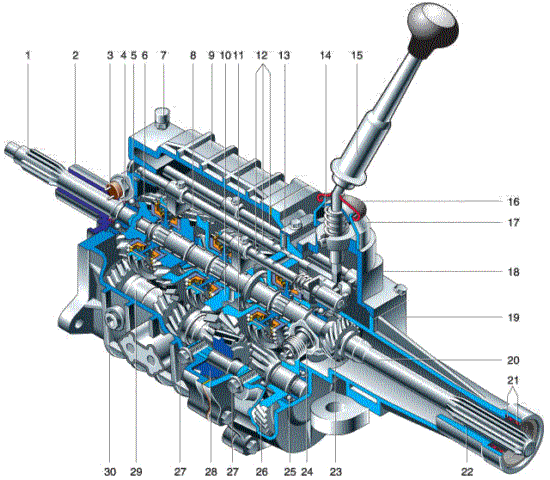
Какая коробка передач лучше – зависит исключительно от условий эксплуатации.
АКПП прекрасно себя зарекомендовала при городской езде. 5-ти или 6-ти ступенчатая, обеспечивающая плавный ход автомобиля, избавляющая водителя от частого переключения передач вовремя езды в вечерних «тянучках». Бывают экземпляры и с восемью ступенями. Но всякий комфорт имеет свою цену. Повышенный расход топлива и медленный разгон автомобиля – умеренная плата за возможность отдохнуть во время пути домой.
Роботизированные коробки предоставляют своим хозяевам практически тот же уровень комфорта. Расход топлива держится на уровне механики, но вот скорость срабатывания оставляет желать лучшего. Изготавливаются роботы на основе современных 5-ти или 6-ти ступенчатых МКП. Прекрасный вариант для городской машины и езды на дальние расстояния по гладкой дороге.
Механическая четырехступенчатая коробка звезд с неба не хватает, но прекрасно выполняет свое основное предназначение. Различные типы 5-ти и 6-ти ступенчатых коробок помотают с непривычки нервы в городе, но прекрасно покажут себя на трассе, поддерживая оптимальный режим работы двигателя.
Что же важнее – комфорт в городе или могучий рывок на трассе, каждый должен решить для себя.
Что такое КПП организации, расшифровка, поиск КПП по ИНН — Контур.Бухгалтерия — СКБ Контур
У многих организаций в реквизитах можно увидеть код причины постановки на учет. В документах КПП идет сразу за ИНН в одной строке «ИНН/КПП». Что это за код, как и где узнать КПП организации, расскажем далее.
Что такое КПП организации
КПП — это набор цифр, дополняющий ИНН. По нему определяют, на основании чего юрлицо поставлено на учет. Включает 9 знаков. Расшифровывается КПП в реквизитах организации так:
- первые две цифры — код региона или области РФ, где стоит на учете фирма;
- вторая пара цифр — номер ИФНС, которая поставила на учет компанию или обособленное подразделение по месту их регистрации, местонахождения недвижимости или транспорта. Также это может быть номер инспекции, которая совершала иные действия по регистрации;
- пятый и шестой знаки — непосредственно причина учета. Для российских организаций доступны значения от 01 до 50, для иностранных компаний — от 51 до 99. В отличие от всех остальных знаков, здесь могут быть и латинские буквы;
- последние цифры в КПП организации — порядковый номер.
 Для российских организаций доступны значения от 01 до 50, для иностранных компаний — от 51 до 99. В отличие от всех остальных знаков, здесь могут быть и латинские буквы;
Для российских организаций доступны значения от 01 до 50, для иностранных компаний — от 51 до 99. В отличие от всех остальных знаков, здесь могут быть и латинские буквы;
По расшифровке можно понять как присваивается КПП организации. Важные моменты — место регистрации и причина.
Что можно узнать по КПП организации
В первую очередь можно узнать налоговую по КПП организации. Для этого достаточно взглянуть на цифры 1-4 — это и есть код ИФНС. Следовательно, по номеру можно узнать регион, где работает юрлицо.
Важную информацию скрывают знаки 5 и 6. Например, комбинация 01 говорит, что КПП присвоен юрлицу при регистрации по местонахождению. Комбинации 06-08 — КПП присвоен по месту нахождения недвижимости. Для крупнейших налогоплательщиков пятая и шестая цифры — 5 и 0 соответственно.
Найти организацию по КПП на сайте налоговой нельзя, потому что это не уникальный номер. Для проверки контрагента используйте ИНН или ОГРН.
Для проверки контрагента используйте ИНН или ОГРН.
Узнать КПП организации по ИНН можно онлайн. Для этого откройте сайт ФНС и получите выписку из ЕГРЮЛ. В ней будет указан нужный номер.
Кому присваивается КПП
Код есть только у организаций. Предприниматели обходятся без него. А кредитные организации часто упускают свой КПП, даже при его наличии. Вот перечень документов, где есть КПП организации:
- налоговых декларациях;
- платежных поручениях;
- счет-фактурах, накладных, УПД и так далее.
КПП всегда рядом с ИНН, но есть ряд отличий. Во-первых, у двух компаний может быть один код, если они стоят на учете в одной ИФНС и по одному основанию. С ИНН такое невозможно.
Во-вторых, КПП не постоянен. Например, код необходимо изменить при смене места регистрации или при становлении крупнейшим налогоплательщиком.
В-третьих, номеров КПП у организации может быть несколько.![]() Самый частый пример — крупнейшие налогоплательщики. Один КПП присвоен при постановке на учет в ИФНС по месту регистрации, второй — при постановке в качестве крупнейшего плательщика налогов. У таких фирм КПП начинается с 99 — это код межрегиональной инспекции по крупнейшим налогоплательщикам.
Самый частый пример — крупнейшие налогоплательщики. Один КПП присвоен при постановке на учет в ИФНС по месту регистрации, второй — при постановке в качестве крупнейшего плательщика налогов. У таких фирм КПП начинается с 99 — это код межрегиональной инспекции по крупнейшим налогоплательщикам.
Регуляция клеточного цикла контрольными точками
1. Митчисон Дж.М. Делящиеся дрожжи Schizosaccharomyces pombe. Биоэссе. 1990;12(4):189–191. [PubMed] [Google Scholar]
2. Mitchison JM, Nurse P. Рост длины клеток у делящихся дрожжей Schizosaccharomyces pombe. Дж. Клеточные науки. 1985; 75: 357–376. [PubMed] [Google Scholar]
3. Митчисон Дж.М. Рост одиночных клеток. I. Schizosaccharomyces pombe. Разрешение ячейки опыта. 1957; 13 (2): 244–262. [PubMed] [Google Scholar]
4. Фантес П.А. Контроль размера клеток и времени цикла у Schizosaccharomyces pombe. Дж. Клеточные науки. 1977;24(51):51–67. [PubMed] [Google Scholar]
5. Rao PN, Johnson RT. Слияние клеток млекопитающих. IV. Регуляция образования хромосом из интерфазных ядер различными химическими соединениями. J Cell Physiol. 1971;78(2):217–223. [PubMed] [Google Scholar]
IV. Регуляция образования хромосом из интерфазных ядер различными химическими соединениями. J Cell Physiol. 1971;78(2):217–223. [PubMed] [Google Scholar]
6. Джонсон Р.Т., Рао П.Н., Хьюз Х.Д. Слияние клеток млекопитающих. 3. Клеточный индуктор преждевременной конденсации хромосом HeLa, активный в клетках различных видов животных. J Cell Physiol. 1970;76(2):151–157. [PubMed] [Академия Google]
7. Джонсон Р.Т., Рао П.Н. Слияние клеток млекопитающих: индукция преждевременной конденсации хромосом в интерфазных ядрах. Природа. 1970; 226(5247):717–722. [PubMed] [Google Scholar]
8. Rao PN, Johnson RT. Слияние клеток млекопитающих: исследования регуляции синтеза ДНК и митоза. Природа. 1970; 225(5228):159–164. [PubMed] [Google Scholar]
9. Маллер Дж.Л. MPF и контроль клеточного цикла. Adv Second Messenger Phosphoprotein Res. 1990; 24: 323–328. [PubMed] [Google Scholar]
10. Масуи Ю. От созревания ооцитов к клеточному циклу in vitro: история открытия фактора, стимулирующего созревание (MPF), и цитостатического фактора (CSF). 2001;69(1): 1–17. [PubMed] [Google Scholar]
2001;69(1): 1–17. [PubMed] [Google Scholar]
11. Weinert T, Hartwell L. Ген RAD9 контролирует реакцию клеточного цикла на повреждение ДНК у Saccharomyces cerevisiae. Наука. 1988; 241:317–322. [PubMed] [Google Scholar]
12. Hartwell L, Weinert T. Контрольные точки: элементы управления, которые обеспечивают порядок событий клеточного цикла. Наука. 1989; 246: 629–634. [PubMed] [Google Scholar]
13. O’Connell MJ, Cimprich KA. Контрольные точки повреждений G2: что включается? Дж. Клеточные науки. 2005; 118 (часть 1): 1–6. [PubMed] [Академия Google]
14. О’Коннелл М.Дж., Уолворт, Северная Каролина, Карр А.М. Контрольная точка повреждения ДНК фазы G2. Тенденции клеточной биологии. 2000;10(7):296–303. [PubMed] [Google Scholar]
15. Tapia-Alveal C, Calonge TM, O’Connell MJ. Регуляция Чк1. Сотовый отдел 2009;4(1):8. [Бесплатная статья PMC] [PubMed] [Google Scholar]
16. Kuntz K, O’Connell MJ. Контрольная точка повреждения ДНК G(2): может ли этот древний регулятор быть ахиллесовой пятой рака? Рак Биол Тер. 2009;8(15):1433–1439. [PubMed] [Академия Google]
2009;8(15):1433–1439. [PubMed] [Академия Google]
17. Хойт М.А., Тотис Л., Робертс Б.Т. S.cerevisiae генов, необходимых для остановки клеточного цикла в ответ на потерю функции микротрубочек. Клетка. 1991; 66: 507–517. [PubMed] [Google Scholar]
18. Ли Р., Мюррей А.В. Обратная связь митоза у почкующихся дрожжей. Клетка. 1991; 66: 519–531. [PubMed] [Google Scholar]
19. Лара-Гонсалес П., Вестхорп Ф.Г., Тейлор С.С. КПП сборки шпинделя. Карр Биол. 2012;22(22):R966–R980. [PubMed] [Google Scholar]
20. Goodarzi AA, Jeggo PA. Репарационные и сигнальные реакции на двухцепочечные разрывы ДНК. Ад Генет. 2013; 82:1–45. [PubMed] [Академия Google]
21. Grallert B, Boye E. Несколько аспектов контрольно-пропускного пункта intra-S. Клеточный цикл. 2008;7(15):2315–2320. [PubMed] [Google Scholar]
22. Эррико А., Костанцо В. Механизмы защиты репликационной вилки: гарантия стабильности генома. Crit Rev Biochem Mol Biol. 2012;47(3):222–235. [PubMed] [Google Scholar]
23. Ламберт С., Карр А.М. Репликационный стресс и перестройки генома: уроки дрожжевых моделей. Curr Opin Genet Dev. 2013;23(2):132–139. [PubMed] [Академия Google]
Ламберт С., Карр А.М. Репликационный стресс и перестройки генома: уроки дрожжевых моделей. Curr Opin Genet Dev. 2013;23(2):132–139. [PubMed] [Академия Google]
24. Ламберт С., Карр А.М. Ответы контрольных точек на барьеры вилки репликации. Биохимия. 2005;87(7):591–602. [PubMed] [Google Scholar]
25. Giono LE, Manfredi JJ. Опухолевой супрессор p53 участвует в нескольких контрольных точках клеточного цикла. J Cell Physiol. 2006;209(1):13–20. [PubMed] [Google Scholar]
26. Килландер Д., Зеттерберг А. Количественное цитохимическое исследование взаимосвязи между клеточной массой и инициацией синтеза ДНК в фибробластах мыши in vitro. Разрешение ячейки опыта. 1965;40(1):12–20. [PubMed] [Google Scholar]
27. Килландер Д., Зеттерберг А. Количественные цитохимические исследования межфазного роста. I. Определение ДНК, РНК и массового содержания возрастных фибробластов мыши in vitro и межклеточной изменчивости во времени генерации. Разрешение ячейки опыта. 1965; 38: 272–284.![]() [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
28. Cross F. DAF1, мутантный ген, влияющий на контроль размера, блокировку феромонов и кинетику клеточного цикла S. cerevisiae. Мол Селл Биол. 1988; 8: 4675–4684. [Бесплатная статья PMC] [PubMed] [Google Scholar]
29. Нэш Р., Токива Г., Ананд С., Эриксон К., Футчер А.Б. Ген WHI1+ Saccharomyces cerevisiae привязывает клеточное деление к размеру клетки и является гомологом циклина. EMBO J. 1988;7(13):4335–4346. [Бесплатная статья PMC] [PubMed] [Google Scholar]
30. Морено С., Медсестра П., Рассел П. Регуляция митоза путем циклического накопления митотического индуктора p80cdc25 у делящихся дрожжей. Природа. 1990;344(6266):549–552. [PubMed] [Google Scholar]
31. Медсестра П. Генетический контроль размера клеток при делении клеток у дрожжей. Природа. 1975;256:547–551. [PubMed] [Google Scholar]
32. Йоргенсен П., Нишикава Дж. Л., Брейткройц Б. Дж., Тайерс М. Систематическая идентификация путей, которые связывают рост и деление клеток у дрожжей. Наука. 2002;297(5580):395–400. [PubMed] [Google Scholar]
Наука. 2002;297(5580):395–400. [PubMed] [Google Scholar]
33. Martin SG, Berthelot-Grosjean M. Полярные градиенты киназы семейства DYRK Pom1 связывают длину клетки с клеточным циклом. Природа. 2009;459(7248):852–856. [PubMed] [Google Scholar]
34. Moseley JB, Mayeux A, Paoletti A, Nurse P. Пространственный градиент координирует размер клеток и митотический вход у делящихся дрожжей. Природа. 2009 г.;459(7248):857–860. [PubMed] [Google Scholar]
35. Аль-Ходаири Ф., Карр А.М. Мутанты репарации ДНК, определяющие пути контрольных точек G2 у Schizosaccharomyces pombe. EMBO J. 1992; 11 (4): 1343–1350. [Бесплатная статья PMC] [PubMed] [Google Scholar]
36. Al-Khodairy F, Fotou E, Sheldrick KS, Griffiths DJF, Lehman AR, Carr AM. Идентификация и характеристика новых элементов, участвующих в контроле контрольных точек и обратной связи у делящихся дрожжей. Мол Биол Селл. 1994; 5: 147–160. [Бесплатная статья PMC] [PubMed] [Google Scholar]
37. Enoch T, Carr A, Nurse P. Гены делящихся дрожжей, участвующие в соединении митоза с завершением репликации ДНК. Гены Дев. 1992;6:2035–2046. [PubMed] [Google Scholar]
Гены делящихся дрожжей, участвующие в соединении митоза с завершением репликации ДНК. Гены Дев. 1992;6:2035–2046. [PubMed] [Google Scholar]
38. Enoch T, Gould K, Nurse P. Cold Spring Harbour Symp Quant Biol. Том. 56. Лабораторный пресс CSH; 1991. Контроль митотических контрольных точек у делящихся дрожжей; стр. 409–416. [PubMed] [Google Scholar]
39. Ван Б., Мацуока С., Карпентер П.Б., Элледж С.Дж. 53BP1, медиатор контрольной точки повреждения ДНК. Наука. 2002;298 (5597): 1435–1438. [PubMed] [Google Scholar]
40. Голдберг М., Штуки М., Фальк Дж., Д’Амур Д., Рахман Д., Паппин Д., Бартек Дж., Джексон С.П. MDC1 необходим для контрольной точки повреждения ДНК внутри S-фазы. Природа. 2003; 421 (6926): 952–956. [PubMed] [Google Scholar]
41. Stewart GS, Wang B, Bignell CR, Taylor AM, Elledge SJ. MDC1 является медиатором контрольной точки повреждения ДНК млекопитающих. Природа. 2003;421(6926):961–966. [PubMed] [Google Scholar]
42. Kumagai A, Dunphy WG. Claspin, новый белок, необходимый для активации Chk1 во время ответа контрольной точки репликации ДНК в экстрактах яиц Xenopus. Мол Ячейка. 2000;6(4):839–849. [PubMed] [Google Scholar]
Claspin, новый белок, необходимый для активации Chk1 во время ответа контрольной точки репликации ДНК в экстрактах яиц Xenopus. Мол Ячейка. 2000;6(4):839–849. [PubMed] [Google Scholar]
43. Latif C, Elzen NR, O’Connell MJ. Поддержание контрольной точки повреждения ДНК за счет устойчивой активности Chk1. Дж. Клеточные науки. 2004; 117 (часть 16): 3489–3498. [PubMed] [Google Scholar]
44. MacDougall CA, Byun TS, Van C, Yee MC, Cimprich KA. Структурные детерминанты активации контрольных точек. Гены Дев. 2007;21(8):898–903. [Бесплатная статья PMC] [PubMed] [Google Scholar]
45. Lu X, Nannenga B, Donehower LA. PPM1D дефосфорилирует Chk1 и p53 и устраняет контрольные точки клеточного цикла. Гены Дев. 2005;19(10): 1162–1174. [Бесплатная статья PMC] [PubMed] [Google Scholar]
46. den Elzen N, Kosoy A, Christopoulos H, O’Connell MJ. Сопротивление аресту: восстановление после ареста контрольной точки посредством дефосфорилирования Chk1 с помощью PP1. Клеточный цикл. 2004;3(5):529–533. [PubMed] [Google Scholar]
2004;3(5):529–533. [PubMed] [Google Scholar]
47. den Elzen NR, O’Connell MJ. Восстановление после остановки контрольной точки повреждения ДНК за счет ингибирования Chk1, опосредованного PP1. Эмбо Дж. 2004; 23(4):908–918. [Бесплатная статья PMC] [PubMed] [Google Scholar]
48. Allen JB, Zhou Z, Siede W, Friedberg E, Elledge S. The SAD1/Rad53 протеинкиназа контролирует множественные контрольные точки и транскрипцию, индуцированную повреждением ДНК, у дрожжей. Гены Дев. 1994; 8: 2416–2428. [PubMed] [Google Scholar]
49. Sun Z, Hsiao J, Fay DS, Stern DF. Домен FHA Rad53 связан с фосфорилированным Rad9 в контрольной точке повреждения ДНК. Наука. 1998;281(5374):272–274. [см. комментарии] [PubMed] [Google Scholar]
50. Cohen-Fix O, Koshland D. Ингибитор анафазы Saccharomyces cerevisiae Pds1p является мишенью пути контрольной точки повреждения ДНК. Proc Natl Acad Sci U S A. 1997;94(26):14361–14366. [Бесплатная статья PMC] [PubMed] [Google Scholar]
51. Rieder CL, Cole RW. Вступление в митоз в соматических клетках позвоночных охраняется контрольной точкой повреждения хромосом, которая обращает клеточный цикл при запуске во время ранней, но не поздней профазы. Джей Селл Биол. 1998;142(4):1013–1022. [Статья бесплатно PMC] [PubMed] [Google Scholar]
Rieder CL, Cole RW. Вступление в митоз в соматических клетках позвоночных охраняется контрольной точкой повреждения хромосом, которая обращает клеточный цикл при запуске во время ранней, но не поздней профазы. Джей Селл Биол. 1998;142(4):1013–1022. [Статья бесплатно PMC] [PubMed] [Google Scholar]
52. Zhou Z, Elledge S. DUN1 кодирует протеинкиназу, которая контролирует реакцию на повреждение ДНК у дрожжей. Клетка. 1993;75:1119–1127. [PubMed] [Google Scholar]
53. Carvajal LA, Hamard PJ, Tonnessen C, Manfredi JJ. E2F7, новая мишень, активируется р53 и опосредует зависимую от повреждения ДНК репрессию транскрипции. Гены Дев. 2012;26(14):1533–1545. [Бесплатная статья PMC] [PubMed] [Google Scholar]
54. St Clair S, Giono L, Varmeh-Ziaie S, Resnick-Silverman L, Liu WJ, Padi A, Dastidar J, DaCosta A, Mattia M, Manfredi JJ . Индуцированное повреждением ДНК подавление Cdc25C опосредуется p53 посредством двух независимых механизмов: один включает прямое связывание с промотором cdc25C. Мол Ячейка. 2004;16(5):725–736. [PubMed] [Академия Google]
Мол Ячейка. 2004;16(5):725–736. [PubMed] [Академия Google]
55. Карвахаль Л.А., Манфреди Дж.Дж. Еще одна развилка на пути — решение о жизни или смерти принимает супрессор опухоли p53. EMBO Rep. 2013;14(5):414–421. [Бесплатная статья PMC] [PubMed] [Google Scholar]
56. Nordstrom W, Abrams JM. Предки-хранители: муха p53 и индуцируемый повреждением апоптоз. Смерть клеток 2000;7(11):1035–1038. [PubMed] [Google Scholar]
57. Nishitani H, Lygerou Z. Лицензирование репликации ДНК. Фронт биосай. 2004; 9: 2115–2132. [PubMed] [Google Scholar]
58. Xu YJ, Davenport M, Kelly TJ. Двухстадийный механизм активации киназы контрольной точки репликации ДНК Cds1 у делящихся дрожжей. Гены Дев. 2006;20(8):990–1003. [Бесплатная статья PMC] [PubMed] [Google Scholar]
59. Bailis JM, Luche DD, Hunter T, Forsburg SL. Поддерживающие белки минихромосомы взаимодействуют с контрольными точками и белками рекомбинации, способствуя стабильности генома s-фазы. Мол Селл Биол. 2008;28(5):1724–1738. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
60. Stead BE, Brandl CJ, Sandre MK, Davey MJ. Фосфорилирование Mcm2 и ответ на репликативный стресс. БМС Генет. 2012;13:36. [Бесплатная статья PMC] [PubMed] [Google Scholar]
61. Sweeney FD, Yang F, Chi A, Shabanowitz J, Hunt DF, Durocher D. Saccharomyces cerevisiae Rad9 действует как адаптер Mec1, позволяющий активировать Rad53. Карр Биол. 2005;15(15):1364–1375. [PubMed] [Google Scholar]
62. Lee KY, Myung K. Модификации PCNA для регуляции путей восстановления после репликации. Мол клетки. 2008;26(1):5–11. [Статья бесплатно PMC] [PubMed] [Google Scholar]
63. Rhind N, Furnari B, Russell P. Фосфорилирование тирозина Cdc2 необходимо для контрольной точки повреждения ДНК у делящихся дрожжей. Гены Дев. 1997;11:504–511. [PubMed] [Google Scholar]
64. Линдси Х.Д., Гриффитс Д.Дж.Ф., Эдвардс Р.Дж., Кристенсен П.У., Мюррей Дж.М., Осман Ф., Уолворт Н., Карр А.М. Специфичная для S-фазы активация киназы Cds1 определяет подпуть ответа контрольной точки у Schizosaccharomyces pombe. Гены Дев. 1998; 12: 382–395. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Гены Дев. 1998; 12: 382–395. [Бесплатная статья PMC] [PubMed] [Google Scholar]
65. Zeng Y, Forbes KC, Wu Z, Moreno S, Piwnica-Worms H, Enoch T. Контрольная точка репликации требует фосфорилирования фосфатазы cdc25 с помощью cds1 или chk1. Природа. 1998;395:507–510. [PubMed] [Google Scholar]
66. Furnari B, Blasina A, Boddy MN, McGowan CH, Russell P. Cdc25 ингибируется in vivo и in vitro киназами контрольных точек Cds1 и Chk1. Мол Биол Селл. 1999;10(4):833–845. [Бесплатная статья PMC] [PubMed] [Google Scholar]
67. Лундгрен К., Уолворт Н., Бухер Р., Дембски М., Киршнер М., Бич Д. mik1 и wee1 взаимодействуют в ингибирующем фосфорилировании тирозина cdc2. Клетка. 1991;64(6):1111–1122. [PubMed] [Google Scholar]
68. Нишитани Х., медсестра П. стр. 65 cdc18 играет важную роль, контролируя инициацию репликации ДНК у делящихся дрожжей. Клетка. 1995; 83: 397–405. [PubMed] [Google Scholar]
69. Морено С., медсестра П. Регуляция прохождения фазы G1 клеточного цикла с помощью гена rum1 + . Природа. 1994; 367: 236–242. [PubMed] [Google Scholar]
Природа. 1994; 367: 236–242. [PubMed] [Google Scholar]
70. Hayles J, Fisher D, Woollard A, Nurse P. Временной порядок S-фазы и митоза у делящихся дрожжей определяется состоянием p34 cdc2 /митотический В-циклиновый комплекс. Клетка. 1994; 78: 813–822. [PubMed] [Google Scholar]
71. O’Connell MJ, Nurse P. Как клетки узнают, что они находятся в G1 или G2. Curr Opin Cell Biol. 1994;6(6):867–871. [PubMed] [Google Scholar]
72. Виттманн Т., Хайман А., Десаи А. Веретено: динамическая сборка микротрубочек и моторов. Nat Cell Biol. 2001;3(1):E28–E34. [PubMed] [Google Scholar]
73. McLean JR, Chaix D, Ohi MD, Gould KL. Состояние APC/C: организация, функции и структура. Crit Rev Biochem Mol Biol. 2011;46(2):118–136. [Бесплатная статья PMC] [PubMed] [Google Scholar]
74. О’Коннелл М.Дж., Криен М.Дж., Хантер Т. Никогда не говори никогда. Родственные NIMA протеинкиназы в митотическом контроле. Тенденции клеточной биологии. 2003;13(5):221–228. [PubMed] [Google Scholar]
75. Малумбрес М., Барбацид М. Киназы клеточного цикла при раке. Curr Opin Genet Dev. 2007;17(1):60–65. [PubMed] [Google Scholar]
Малумбрес М., Барбацид М. Киназы клеточного цикла при раке. Curr Opin Genet Dev. 2007;17(1):60–65. [PubMed] [Google Scholar]
76. Koniaras K, Cuddihy AR, Christopolous H, Hogg A, O’Connell MJ. Ингибирование Chk1-зависимой контрольной точки повреждения ДНК G2 вызывает радиосенсибилизацию мутантных клеток p53 человека. Онкоген. 2001; 20:7453–7463. [PubMed] [Академия Google]
77. Могери-Сакка М., Бартуччи М., Де Мария Р. Ингибиторы контрольной точки киназы 1 для усиления системной противоопухолевой терапии. Cancer Treat Rev. 2013;39(5):525–533. [PubMed] [Google Scholar]
78. Stathis A, Oza A. Ориентация на Wee1-подобную протеинкиназу для лечения рака. Перспектива новостей о наркотиках. 2010;23(7):425–429. [PubMed] [Google Scholar]
79. Walworth N, Davey S, Beach D. Протеинкиназа делящихся дрожжей chk1 связывает путь контрольной точки rad с cdc2 . Природа. 1993;363:368–371. [PubMed] [Google Scholar]
80. Liu Q, Guntuku S, Cui XS, Matsuoka S, Cortez D, Tamai K, Luo G, Carattini-Rivera S, DeMayo F, Bradley A, Donehower LA, Elledge SJ.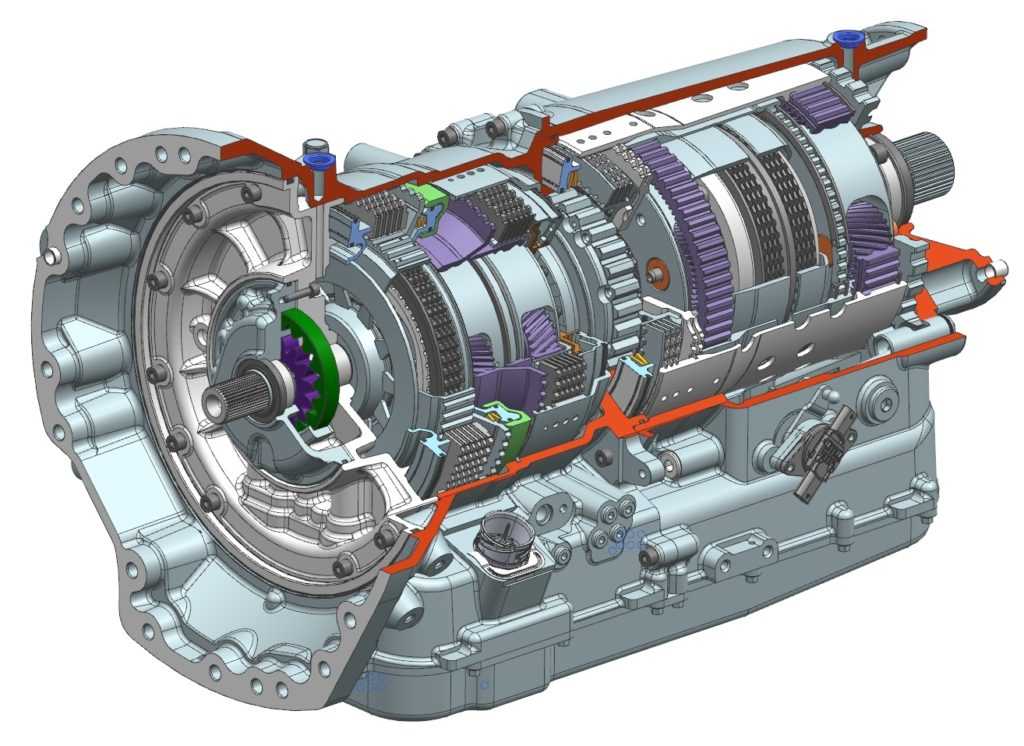 Chk1 является важной киназой, которая регулируется Atr и необходима для контрольной точки повреждения ДНК G(2)/M. Гены Дев. 2000;14(12):1448–1459. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Chk1 является важной киназой, которая регулируется Atr и необходима для контрольной точки повреждения ДНК G(2)/M. Гены Дев. 2000;14(12):1448–1459. [Бесплатная статья PMC] [PubMed] [Google Scholar]
81. Фогарти П., Кэмпбелл С.Д., Абу-Шумайс Р., Фалле Б.С., Ю К.Р., Уй Г.Л., Голдберг М.Л., Салливан В. Ген винограда дрозофилы связан с контрольной точкой ген chk1/rad27 и необходим для точности позднего синцитиального деления. Карр Биол. 1997;7(6):418–426. [PubMed] [Google Scholar]
82. Гриноу К.Р., Кларк А.Р., Джонс Р.Х. Дефицит Chk1 в тонком кишечнике мышей приводит к p53-независимой гибели крипт и последующей кишечной компенсации. Онкоген. 2009;28(11):1443–1453. [Бесплатная статья PMC] [PubMed] [Google Scholar]
83. Smith J, Larue L, Gillespie DA. Chk1 необходим для развития мышиных эпидермальных меланоцитов. Пигментно-клеточная меланома Res. 2013;26(4):580–585. [PubMed] [Google Scholar]
84. Lam MH, Liu Q, Elledge SJ, Rosen JM. Chk1 гаплонедостаточен для многих функций, важных для подавления опухоли. Раковая клетка. 2004;6(1):45–59.. [PubMed] [Google Scholar]
Раковая клетка. 2004;6(1):45–59.. [PubMed] [Google Scholar]
85. Петерманн Э., Майя-Мендоза А., Захос Г., Гиллеспи Д.А., Джексон Д.А., Калдекотт К.В. Требование Chk1 для высоких глобальных скоростей прогрессирования репликационной вилки во время S-фазы нормальных позвоночных. Мол Селл Биол. 2006;26(8):3319–3326. [Бесплатная статья PMC] [PubMed] [Google Scholar]
86. Zachos G, Rainey MD, Gillespie DA. Chk1-дефицитные опухолевые клетки жизнеспособны, но имеют множественные дефекты контрольных точек и выживаемости. EMBO J. 2003;22(3):713–723. [Бесплатная статья PMC] [PubMed] [Google Scholar]
87. Копс Г.Дж., Уивер Б.А., Кливленд Д.В. На пути к раку: анеуплоидия и митотическая контрольная точка. Нат Рев Рак. 2005;5(10):773–785. [PubMed] [Google Scholar]
Клеточный цикл, его фазы и контрольные точки — обзор
Клеточный цикл и его фазы являются одним из центральных понятий клеточной биологии. Он включает в себя серию четко определенных событий, через которые проходит клетка, чтобы скопировать свое генетическое содержимое и разделить на две идентичные дочерние клетки. Это деление клеток является основной особенностью жизни. Все эукариотические клетки, то есть клетки с ядрами, проходят клеточный цикл, чтобы делиться. Несмотря на то, что в некоторых аспектах клетки животных, растительных клеток и грибов различаются, общая процедура для них одинакова.
Это деление клеток является основной особенностью жизни. Все эукариотические клетки, то есть клетки с ядрами, проходят клеточный цикл, чтобы делиться. Несмотря на то, что в некоторых аспектах клетки животных, растительных клеток и грибов различаются, общая процедура для них одинакова.
Основные фазы клеточного цикла
Клеточный цикл обычно делится на четыре основные фазы. Первые три фазы часто называют интерфазой, и они подготавливают клетку к четвертой фазе — митозу.
Схематическое изображение клеточного цикла с указанием основных фаз и контрольных точек.
G
1 фаза – рост клеток
Клетка увеличивается в размерах и увеличивает производство мРНК, белка и клеточных органелл, чтобы подготовиться к более поздним стадиям.
S-фаза – синтез ДНК
Производство мРНК и белка снижается, и клетка фокусируется на репликации всей своей ДНК, создавая по одной копии каждой хромосомы. Две идентичные копии, называемые сестринскими хроматидами, прикрепляются друг к другу в точке, называемой центромерой.
Классическое изображение хромосомы. ДНК была реплицирована, поэтому хромосома состоит из двух идентичных сестринских хроматид, соединенных центромерой.
Г
2 9Фаза 0204 – подготовка к делению клеток Клетка продолжает расти и синтезировать белки и органеллы, необходимые для последней фазы. Ядерная мембрана разрывается, освобождая хромосомы. Сестринские хроматиды разделяются и образуют два идентичных ядра, оттянутых к противоположным сторонам клетки. Затем он разделяет клеточную мембрану и цитоплазму на две равные части, разделяя между ними ДНК и клеточные органеллы. Были созданы две генетически идентичные дочерние клетки. Клетка во время М-фазы. (1) Ядерная мембрана разрушается, освобождая хромосомы. (2) Хромосомы выстраиваются в линию и прикрепляются к микротрубочкам веретена. (3) Хромосомы разрываются на центромере, и микротрубочки тянут сестринские хроматиды к противоположным сторонам клетки. 0 фаза После того, как клетка завершила М-фазу, обе дочерние клетки входят в G 1 снова фаза. Отсюда они могут либо продолжить клеточный цикл и снова делиться, либо выйти из него и перестать делиться. Клетка, выходящая из клеточного цикла, переходит в фазу G 0 , где она все еще полностью функциональна, но больше не делится. Некоторые клетки никогда больше не покидают фазу G 0 , как и большинство нейронных клеток, тогда как другие могут повторно войти в фазу G 1 и при необходимости продолжить клеточный цикл. Как и большинство эпителиальных клеток, некоторые клетки никогда не проникают в G 0 , но продолжают делиться на протяжении всей жизни. Чтобы убедиться, что каждая фаза клеточного цикла выполняется правильно, за основными этапами следует контрольная точка. Клеточный цикл включает несколько контрольных точек, основными из которых являются G 1 , G 2 и М КПП. 1 Контрольный пункт – проверка размера и питательных веществ Эта контрольная точка, также называемая контрольной точкой ограничения, имеет место между G 1 и фазой S. Клетка проверяет, достаточно ли она велика для деления, не повреждена ли ее ДНК и есть ли достаточный доступ к питательным веществам и стимулирующим факторам роста. Если он проходит контрольную точку G 1 , он необратимо завершает весь клеточный цикл и делится. Если нет, он пытается исправить то, что не удается, или войти в G 0 фазы и ждать более благоприятных условий. 2 контрольно-пропускной пункт – контроль качества ДНК После второй фазы роста клетка проверяет, полностью ли и правильно реплицировалась ДНК во время S-фазы. В случае успеха он переходит к фазе М, а в случае неудачи пытается исправить ошибки. Если клетка не может восстановить ДНК, она подвергается апоптозу. Это предотвращает передачу поврежденной ДНК дочерним клеткам. Эта контрольная точка происходит во время M-фазы. Его также называют контрольной точкой веретена, потому что клетка проверяет, правильно ли все сестринские хроматиды прикреплены к микротрубочкам веретена, которые их разделяют. Если нет, клетка приостанавливает митоз до тех пор, пока все сестринские хроматиды не будут прикреплены должным образом. и рост клеток Клеточная пролиферация — это термин, тесно связанный с клеточным циклом, означающий процесс, который увеличивает количество клеток за счет клеточного деления и роста клеток . М-фаза – образование дочерних клеток путем митоза
 (4) Образуются два новых полных ядра, и клетка может делиться на две дочерние клетки.
(4) Образуются два новых полных ядра, и клетка может делиться на две дочерние клетки. Выход из клеточного цикла – G
Контрольные точки клеточного цикла
 Это механизм контроля, в котором должны быть выполнены определенные условия, прежде чем клетка сможет перейти к следующему шагу. Сбой контрольной точки вызывает остановку клеточного цикла, не позволяя ячейке продолжать работу до тех пор, пока она не решит проблемы.
Это механизм контроля, в котором должны быть выполнены определенные условия, прежде чем клетка сможет перейти к следующему шагу. Сбой контрольной точки вызывает остановку клеточного цикла, не позволяя ячейке продолжать работу до тех пор, пока она не решит проблемы. G

G
КПП М — проверка установки ядерного дивизиона
Пролиферация клеток – деление клеток
